Remplacer progressivement les forêts françaises par des plantations d’espèces censées être plus résilientes face au changement climatique. L’idée fait du chemin. Mais ce remède ne risque-t-il pas d’être pire que le mal ? Un décryptage des scientifiques Xavier Morin, chercheur en écologie des forêts au CNRS, Joannès Guillemot, chercheur en physiologie des forêts au Cirad, Max Bruciamacchie, enseignant-chercheur à AgroParisTech, Nicolas Martin, chargé de recherche en Écologie, à l'Inrae.
1 milliards d’arbres après les coupes rases
Planter 1 milliard d’arbres en France hexagonale d’ici 2032 et renouveler ainsi 10 % de la forêt française. Ce projet pharaonique annoncé en 2022 par Emmanuel Macron promet d’avoir ainsi des forêts plus résilientes face au changement climatique. Connues pour leur rôle de puits de carbone, les forêts françaises sont de fait de plus en plus mal en point. Elles ont vu, par exemple, ces dix dernières années, leur capacité à stocker du carbone diminuer presque de moitié.
Un constat inquiétant qui invite certains à plébisciter des changements drastiques, tels que l’augmentation du volume de bois coupé chaque année, voire davantage recourir aux coupes rases pour favoriser le remplacement massif d’espèces « condamnées » par des espèces plus résilientes via des plantations.
Mais ce remède radical ne risque-t-il pas d’être pire que le mal ? Certains éléments invitent en tout cas à la prudence, comme celui du manque de fiabilité des modèles qui sont utilisés pour justifier cette option. Ces derniers négligent notamment les capacités de résilience des arbres actuels ou encore la variabilité des réponses entre les arbres ou les stations où ils se trouvent. En tout cas, il est essentiel de questionner la pertinence générale d’une telle stratégie « ultra-interventionniste », que ce soit au niveau écologique ou économique.
Des prédictions souvent très incertaines
La première incertitude qui invite à la prudence concerne le réchauffement à venir. Si l’on se base sur un scénario de réchauffement global de 4 °C ou 5 °C d’ici à 2100, on sait que seul le chêne vert et le pin d’Alep seraient capables de résister au choc dans la plupart des régions de l’hexagone, sauf peut-être à haute altitude. Mais si l’on se projette dans un scénario moins extrême, par exemple à +2.5 °C de plus d’ici à 2100, des solutions plus fines d’adaptation de nos peuplements forestiers s’ouvrent à nous.
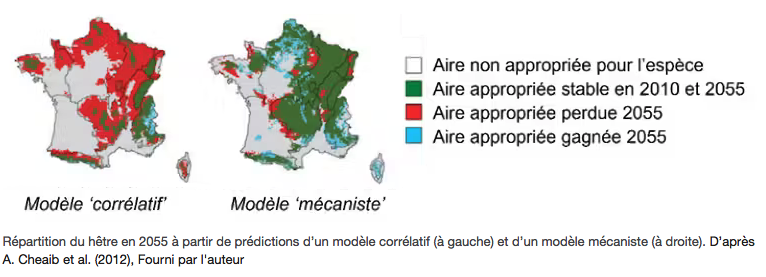
La deuxième incertitude concerne les outils utilisés pour évaluer le devenir des forêts face au réchauffement climatique. La méthode la plus répandue ne repose pas sur la biologie des espèces, comme leur résistance physiologique au stress hydrique ou la réponse de leur croissance face au climat, mais utilise uniquement l’association entre des données spatiales de répartition des essences d’une part et de climat d’autre part.
Ces modèles, dits « corrélatifs », se contentent en effet de faire le lien entre les sites où une espèce est observée et les conditions climatiques de ces sites. Avec une telle approche on peut donc par exemple « prédire » que d’après sa répartition actuelle, le hêtre ne sera présent, au mieux, que dans les sites ayant une température moyenne annuelle comprise entre 4 °C et 13 °C. Ces modèles corrélatifs sont plébiscités depuis la fin des années 1990 car ils sont relativement faciles à mettre en place, et ce pour beaucoup d’espèces. Ce sont donc eux qui sont les plus repris dans les médias et par les décideurs, par exemple le site Nos Forêts demain qui propose des projections des principales espèces européennes.
Mais pourquoi est-ce si problématique ? Parce que, d’une part, ce genre d’approche n’est pas exempte de défauts voire de biais et que, d’autre part, les prédictions faites avec ces outils sont souvent très pessimistes quant à la capacité des espèces à résister au changement climatique.

Crédit : Ollo / iStock
Par exemple, ces modèles corrélatifs ne prennent pas en compte la variabilité individuelle existant d’un arbre à l’autre au sein d’une même espèce. Tout d’abord, des arbres peuvent être localement particulièrement adaptés à leur environnement sous l’effet de la sélection naturelle, c’est ce qu’on appelle l’adaptation locale. Reprenons le cas du hêtre : il y a des populations de hêtres dans le centre de l’Espagne et au sud de la Suède, qui montrent de caractéristiques très différentes que ce soit pour la résistance à la sécheresse ou la sensibilité au gel.
Ensuite, deux arbres d’une même espèce peuvent développer des caractéristiques très différentes selon les conditions dans lesquelles ils vivent, même sans différence génétique : c’est la plasticité phénotypique. Par exemple, un hêtre poussant dans des conditions fraîches et humides, aura développé des feuilles plus larges et nombreuses lui permettant une meilleure croissance. Mais il sera aussi très probablement sensible à des stress hydriques, car ces grandes surfaces foliaires impliquent de plus forte consommation d’eau. À l’inverse, un hêtre ayant grandi dans des stations plus sèches aura tendance à développer des feuilles plus petites, plus épaisses et moins nombreuses, lui permettant de limiter les pertes en eau et de mieux résister aux sécheresses.
Mais, il y a de nombreux autres aspects qui sont ignorés par les modèles corrélatifs – basés seulement sur un lien statistique entre répartition des espèces et climat – et qui biaisent leurs résultats comme : l’utilisation des terres (la forêt n’est pas présente partout où elle le pourrait), la gestion forestière (le forestier impacte fortement les espèces qui sont présentes sur un site donné), l’effet de l’interaction avec d’autres espèces (compétition, herbivorie, parasitisme…), sans parler de problèmes méthodologiques inhérents à cette méthode d’enveloppe climatique.
Des approches alternatives existent pour prédire la répartition future des espèces, qu’on appelle approches mécanistes car davantage basées sur les mécanismes biologiques et écologiques, mais qui sont encore peu diffusées car plus longues à développer. Des comparaisons entre ces différentes méthodes montrent, en moyenne, que les modèles mécanistes prédisent moins d’extinctions locales que les modèles corrélatifs, en particulier pour les arbres. Ceci est illustré pour le hêtre dans l’exemple ci-dessous, où le modèle corrélatif, basé sur une réponse « fixe » de l’espèce, prédit des taux d’extinction plus forts que le modèle mécaniste qui lui inclut notamment la plasticité phénotypique (Fig. X).
Même s’ils ne sont évidemment pas parfaits, les approches basées sur les mécanismes doivent mieux percoler dans les milieux de la gestion, mais en premier lieu il convient d’inciter à davantage de prudence avec les prédictions de modèles corrélatifs et de tempérer leur utilisation pour prendre des décisions opérationnelles.
Mieux coordonner urgence d’action et temporalité des cycles forestiers
Autre inquiétude légitime que peut soulever l’approche très interventionniste qui viserait à remplacer des forêts entières actuelles par des peuplements de « nouvelles » espèces : celle d’une confusion entre le cycle de développement d’un arbre et d’une forêt.
Pour cela, replaçons-nous dans la temporalité de ces projets de remplacement des forêts françaises. Ils partent du principe que puisque le changement climatique est rapide, et qu’un peuplement forestier met des décennies à être mature, si l’on doit remplacer les espèces d’arbres, il faut agir très vite pour espérer avoir des forêts (et qui plus est productives) d’ici la fin du siècle.
Le temps de production d’un chêne s’évalue certes en siècles, mais le temps à l’échelle de l’arbre n’est pas celui de la forêt, sauf si la forêt est composée d’une seule espèce d’arbre et avec des troncs de taille uniforme. Ce n’est généralement pas le cas. Sous nos latitudes les forêts sont souvent mélangées et si ce n’est pas le cas, elles sont constituées de troncs de taille variée. Dans ce cas, le temps mis pour récupérer le capital de départ tout en ayant in fine conservé ce capital, est très souvent inférieur à trente-cinq ans, parfois même moins de vingt ans, et même dix ans dans des cas exceptionnels. Ce concept permet de relativiser l’urgence des interventions, sans pour autant minimiser l’impact des aléas climatiques, et pourrait être utilisé pour organiser et classer les forêts en fonction de leur capacité à se reconstituer.

Intervenir drastiquement, c’est annihiler toute chance de résilience naturelle
Enfin, miser principalement sur le remplacement des arbres existants c’est aussi se priver des opportunités de résilience naturelle que l’on peut observer localement. Cette résilience peut changer grandement pour une même espèce en fonction des conditions stationnelles (exposition, pente, profondeur et qualité des sols…), du type de peuplement (pur ou en mélanges d’essences, en plantation ou issu de régénération naturelle). Il a par exemple été largement montré que le mélange d’essences peut agir comme une assurance contre les changements : les peuplements mélangés peuvent notamment s’avérer plus résilients aux évènements de sécheresse et aux ravageurs que les peuplements purs, et à ce à tous les stades de développement.
De plus, comme évoqué plus haut, chaque arbre a de fait une capacité propre plus ou moins grande de résilience. Ainsi, couper à blanc une forêt entière au motif que seul un certain nombre d’arbres sont dépérissants conduit potentiellement à éliminer des arbres qui étaient plus résistants. Sachant que la proportion d’arbres dépérissants actuellement retenue pour motiver le renouvellement entier d’un peuplement est faible (20 % des arbres), force est de constater le risque de se priver au passage d’un potentiel génétique crucial pour les conditions futures.
Quant aux plantations censées mettre en place des peuplements résilients dans le futur car composés d’essences « d’avenir », il s’avère que ces derniers sont très fragiles juste après leur installation, notamment du fait de conditions climatiques plus sévères en été. Par exemple, les plantations d’arbres réalisées en 2022 sur le territoire métropolitain ont souffert de taux d’échecs de près de 40 % après seulement quelques mois.
Au passage, en rasant une forêt on génère également de nombreux effets néfastes : l’écosystème cesse d’être un puits de carbone, devenant une source pour plusieurs années voire décennies, et la coupe rase mène nécessairement à une perte de biodiversité, au tassement des sols réduisant leur fertilité, ainsi qu’à de forts impacts sociétaux.
Comme rappelé récemment par la communauté scientifique, les bénéfices de l’intervention massive semblent donc, a minima, peu évidents face au risque de se priver de la résilience naturelle. Cette dernière pourrait être mieux favorisée par d’autres types d’interventions, plus « doux » vis-à-vis de l’écosystème, par exemple l’enrichissement sous couvert ou la sylviculture mélangée à couvert continu. L’enrichissement sous couvert est une technique de gestion forestière qui consiste à introduire de nouvelles espèces sous le couvert d’une forêt déjà existante. La sylviculture mélangée à couvert continu est une approche de gestion forestière qui combine deux éléments essentiels : la diversité des espèces d’arbres (sylviculture mélangée) et le maintien d’une couvert végétal continu de la forêt.
Cette méthode vise à maintenir une forêt stable et productive tout en préservant un couvert forestier constant. Plus généralement, des recherches sur ces sujets sont actuellement en cours, et devraient vite apporter des éléments concrets sur les pistes d’adaptation. Un article scientifique initialement publié sur le site The Conversation, reproduit ici à la demande de ses auteurs.
Un autre monde est possible. Tout comme vivre en harmonie avec le reste du Vivant. Notre équipe de journalistes œuvre partout en France et en Europe pour mettre en lumière celles et ceux qui incarnent leur utopie. Nous vous offrons au quotidien des articles en accès libre car nous estimons que l’information doit être gratuite à tou.te.s. Si vous souhaitez nous soutenir, la vente de nos livres financent notre liberté.


